De kunst van het ontwijken van roofvogels
In de natuur lopen dieren het gevaar ten prooi te vallen aan roofdieren die opduiken vanuit elke mogelijke hoek. Te land, ter zee en in de lucht oefent ‘de wet van de sterkste’ een omgevingsdruk uit op de ontwikkeling van het gedrag van verschillende diersoorten. In acute dreigende situaties maken prooidieren gebruik van aangeboren gedragingen die evolutionair gegroeid zijn om gevaar te ontlopen. Zulke aangeboren gedragingen laten hen toe te reageren op verschillende situaties waar ze nog nooit eerder in aanraking mee kwamen. Dit is onder andere van essentieel belang bij het ontwijken van gevaar, maar het speelt ook een belangrijke rol bij de voortplanting en het zoeken naar voedsel. Zo omhult aangeboren gedrag een uiteenlopend gedragsrepertoire dat instaat voor het voortbestaan van de soort.
Onderzoek naar visueel geïnduceerd defensief gedrag
Decennialang wordt reeds het gedrag bestudeerd van uiteenlopende diersoorten waaronder kikkers, kakkerlakken, apen, fruitvliegen, ratten en muizen, waarvan deze laatste het meest populair zijn omwille van hun makkelijk onderhoud en lage kost. Hieruit leerden we dat verschillende diersoorten mogelijks andere defensieve strategieën gebruiken en dat vele dieren er verschillende gedragingen op nahouden afhankelijk van de specifieke situatie. Om het defensief gedrag van deze dieren te bestuderen, werd vroeger gebruik gemaakt van levende roofvogels die in het laboratorium over de studieobjecten vloog. Dankzij de technologische vooruitgang is deze omslachtige aanpak echter niet meer nodig. De dieren worden nu in een gecontroleerde artificiële omgeving gebracht met continue monitoring waarbij de roofvogel vervangen wordt door een virtuele stimulus die op een scherm geprojecteerd wordt. Dit laat toe dat experimenten reproduceerbaar en gecontroleerd uitgevoerd kunnen worden.
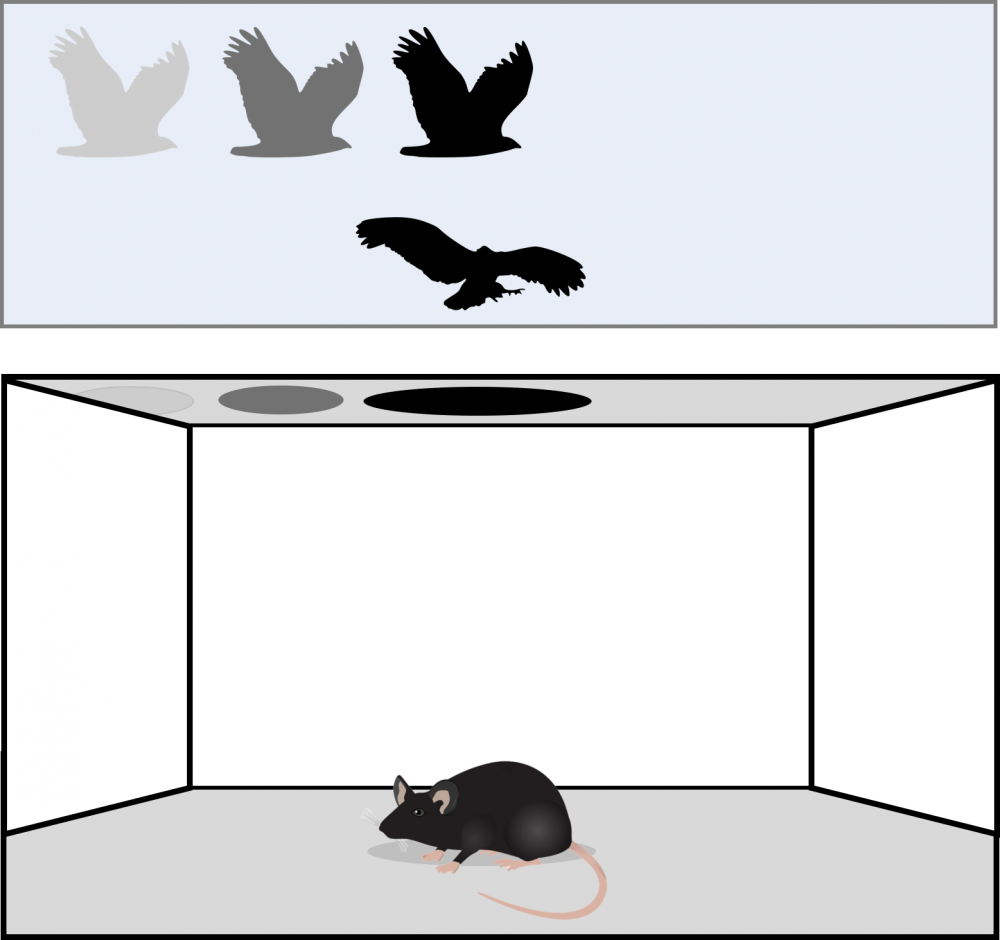
De bevindingen uit zulke testen hebben veel kennis aangereikt binnen de huidige gedragswetenschappen. Zo werd bijvoorbeeld duidelijk dat muizen er twee algemene reactiemechanismen op nahouden wanneer ze in aanraking komen met verschillende dreigingen. Een eerste mechanisme wordt geïnduceerd wanneer de muis een overvliegende roofvogel waarneemt. Om zijn zichtbaarheid voor het roofdier te verlagen, neemt de muis in deze situatie een immobiele houding aan. Echter, wanneer het dreigingsniveau toeneemt, zoals bij een naderende roofvogel, berusten muizen op een tweede reactiemechanisme waarbij ze vluchten naar een veilige plek. Deze diversiteit in het defensief gedragsrepertoire doet de vraag rijzen waarom deze dieren anders reageren in verschillende situaties. Het antwoord op deze fundamentele vraag wordt gezocht in de connectiviteit en werking van verschillende hersendelen. Hoewel reeds enkele specifieke neurale pathways werden blootgelegd die betrokken zijn bij deze twee typerende reactiemechanismen, missen we momenteel nog een diepgaand inzicht in deze neurale werking.

Vergelijking van defensief gedrag in Peromyscus knaagdieren
Om nieuwe kennis en nieuwe inzichten te verwerven op het vlak van zowel defensief gedrag als de onderliggende neurale werking werd het voorbije jaar onderzoek verricht op twee nauwverwante diersoorten die nog niet eerder werden geïntroduceerd in dit vakgebied, n.l. de ‘oldfield mouse’ en de ‘deer mouse’. Hoewel deze knaagdiersoorten, die behoren tot het geslacht Peromyscus, vaak bestempeld worden als ‘muizen’, zijn ze meer verwant met hamsters dan met de bekende laboratoriummuis. Ze komen oorspronkelijk voor in Noord-Amerika waar ze er met meer dan 50 andere Peromyscus-soorten de grootste zoogdiergroep vormen. Omwille van hun rijke omvang en grote aantallen vormen deze dieren een belangrijk onderdeel van het ecosysteem en maakten ze doorheen de 20e eeuw vaak deel uit van wetenschappelijke studies binnen de epidemiologie, fysiologie en evolutiewetenschappen. Er werd destijds zo’n aanzienlijke hoeveelheid data verzameld over deze dieren dat ze bestempeld werden als de ‘Drosophila van de Noord-Amerikaanse zoogdierkunde’. Uit zulke eerdere studies is gebleken dat de deer mouse en de oldfield mouse verschillende aangeboren karakteristieke gedragingen vertonen. Bijvoorbeeld, de deer mouse maakt een simpel hol en berust op een polygame voortplanting, terwijl de oldfield mouse een complex hol construeert en een monogame voorplanting kent, wat een zeldzame eigenschap is onder de knaagdieren. Op basis van deze aangeboren verschillen lijken deze twee Peromyscus-soorten interessante studieobjecten binnen het onderzoek naar defensief gedrag. Het drijft de vraag of hun verschillende karaktertrekken ook gereflecteerd worden in een verschil in defensief gedrag, wat vervolgens als basis kan dienen om de neurale verschillen te achterhalen die dit ondersteunen.

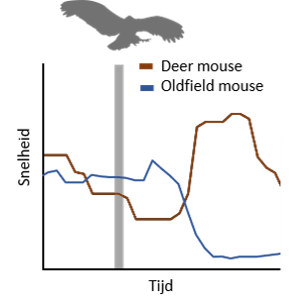
Het onderzoek dat hieraan verbonden was heeft geleid tot een opmerkelijk inzicht in het defensief gedrag van deze Peromyscus-soorten. De resultaten tonen aan dat de nauwverwante dieren een tegenovergesteld gedrag verkiezen in reactie op dezelfde dreiging. Wanneer de oldfield mouse werd blootgesteld aan een virtuele stimulus die een naderende roofvogel imiteerde, nam deze preferentieel een immobiele houding aan. De deer mouse daarentegen vertoonde voornamelijk een vluchtreactie die georiënteerd was in de richting van een schuilplaats. Indien de stimulus reeds verdwenen was vooraleer het dier de schuilplaats binnenging, stopte het voor de ingang. Indien de stimulus langer zichtbaar was, werd de vluchtreactie doorgezet totdat het dier verscholen was. Het merkwaardig verschil tussen immobilisatie en vluchtreactie bij deze twee soorten was enkel zichtbaar in reactie op een naderende roofvogel en is in overeenstemming met hun natuurlijke omgeving. De deer mouse leeft in gebieden met veel vegetatie waarin het zinvol is om te vluchten naar een schuilplaats, terwijl de oldfield mouse in open vlaktes leeft waar zijn zichtbaarheid geminimaliseerd wordt bij een immobiele houding. Een overvliegende roofvogel daarentegen induceerde in beide soorten een immobilisatiereactie die onafhankelijk was van de horizontale snelheid van het roofdier. Deze studie toont bijgevolg aan dat dieren op verschillende defensieve mechanismen kunnen berusten afhankelijk van de soort dreiging en dat nauwverwante soorten onderling verschillend kunnen reageren op hetzelfde gevaar. Deze opmerkelijke resultaten zijn relevant voor neurologisch onderzoek dat neurale elementen tracht te achterhalen die een rol spelen in defensief gedrag. Namelijk, het is waarschijnlijk dat het verschil in reactiestrategieën resulteert van verschillen in geassocieerde neurale pathways die zulk specifiek gedrag ondersteunen. Daarom zouden deze Peromyscus-soorten ons een stap dichter kunnen brengen naar een diepgaand inzicht van de neurale werking bij aangeboren gedrag.
Bibliografie
[1] Evans DA, Stempel AV, Vale R, Branco T. Cognitive control of escape behavior. Trends in Cognitive
Sciences (2019). 23(4):334-348.
[2] Pereira AG, Moita MA. Is there anybody out there? Neural circuits of threat detection in vertebrates.
Current Opinion in Neurobiology (2016). 41:179-187.
[3] Blanchard RJ, Hebert MA, Ferrari P, Palanza P, Figueira R, Blanchard DC, Parmigiani S. Defensive
behaviors in wild and laboratory (Swiss) mice: the mouse defense test battery. Physiology & Bahavior
(1998). 65(2):201-209.
[4] Whishaw IQ, Metz GAS, Kolb B, Pellis SM. Accelerated nervous system development contributes to
behavioral efficiency in the laboratory mouse: a behavioral review and theoretical proposal. Dev
Psychobiol (2001). 39:151-170.
[5] Card G, Dickinson MH. Visually mediated motor planning in the escape response of Drosophila. Curr Biol
(2008). 18(17):1300-1307.
[6] Schiff W, Caviness JA, Gibson JJ. Persistent fear response in rhesus monkeys to the optical stimulus of
“looming”. Science (1962). 136(3520):982-983.
[7] Hu CK, Hoekstra HE. Peromyscus burrowing: a model system for behavioral evolution. Seminars in Cell
& Developmental Biology (2017). 61:107-1114.
[8] Sanes JR, Masland RH. The types of retinal ganglion cells: current status and implications for neural
classification. Annu Rev Neurosci (2015). 18:221-246.
[9] Allen TA, Narayanan NS, Kholodar-Smith DB, Zhao Y, Laubach M, Brown TH. Imaging the spread of
reversible brain inactivations using fluorescent muscimol. J Neurosci Methods (2008). 171(1):30-38.
[10] Karrow K, Masland RH, Physiological clustering of visual channels in the mouse retina. J Neurophysiol
(2011). 105:1516-1530.
[11] Schmidt TM, Chen SK, Hatter S, Intrinsically photosensitive retinal ganglion cells: many subtypes,
diverse functions. Trends Neurosci (2011). 34(11):572-580.
[12] Baden T, Berens P, Franke K, Roson MR, Bethge M, Euler T. The functional diversity of retinal ganglion
cell in the mouse. Nature (2016). 529(7586):345-350.
[13] Morin LP, Studholme KM. Retinofugal projections in the mouse. J Comp Neurol (2014). 522(16):3733-
3753.
[14] Chen SK, Badea TC, Hattar S. Photoentrainment and pupillary light reflex are mediated by distinct
populations of ipRGC. Nature (2012). 476(7358):92-95.
[15] Ito S, Feldheim DA. The mouse superior colliculus: an emerging model for studying circuit formation and
function. Frontiers in Neural Circuits (2018). 12:10.
[16] Schenberg LC, Póvoa RMF, Costa ALP, Caldellas AV, Tufik S, Bittencourt AS. Functional specializations
within the tectum defense systems of the rat. Neuroscience and Biobehavioral Review (2005). 29:1279-
1298.
[17] Byun Haewon, Kwon S, Ahn HJ, Liu H, Forrest D, Demb JB, Kim IJ, Molecular features distinguish ten
neuronal types in the mouse superficial superior colliculus. J Comp Neurol (2016). 524(11):2300-2321.
[18] Shang C, Chen Z, Liu A, Li Y, Zhang J, Qu B, Yan F, Zhang Y, Liu W, Liu Z, Guo X, Li D, Wang Y, Cao
P. Divergent midbrain circuits orchestrate escape and freezing responses to looming stimuli in mice.
Nature Communications (2018). 9:1232.
[19] Reinhard K, Li C, Do Q, Burke E, Heynderickx S, Farrow K. A projection specific logic to sampling visual
inputs in mouse superior colliculus. (2018). doi: https://doi.org/10.1101/272914
[20] Bildstein KL, Forsyth DJ. Effects of dietary dieldrin on behavior of white-footed mice (Peromyscus
leucopus) towards an avian predator. Bull Environm Contam Toxicol (1979). 21:93-97.
[21] Yilmaz M, Meister M. Rapid innate defensive responses of mice to looming visual stimuli. Curr Biol
(2013). 23(20).
[22] Vale R, Evans DA, Branco T. Rapid spatial learning controls instinctive defensive behavior in mice. Curr Biol (2017). 27:1342-1349.
[23] De Franceschi G, Vivattanasarn T, Saleem AB, Solomon SG. Vision guides selection of freeze or flight
defense strategies in mice. Curr Biol (2016). 26:2150-2154.
[24] Evans DA, Stempel AV, Vale R, Ruehle S, Lefler Y, Branco T. A synaptic threshold mechanism for
computing escape decisions. Nature (2018). 558(7711):590-594.
[25] Shorter KR, Owen A, Anderson V, Hall-South AC, Hayford S, Cakora P, Crossland JP, Georgi VRM,
Perkins A, Kelly SJ, Felder MR, Vrana PB. Natural Genetic Variation Underlying Differences in
Peromyscus Repetitive & Social/Aggressive Behaviors. Behav Genet (2014). 44(2):126-135.
[26] Salay LD, Ishiko N, Huberman AD. A midline thalamic circuit determining reactions to visual threat.
Nature (2018). 557(7704):183-189.
[27] Dewey MJ, Dawson WD. Deer mice: “The Drosophila of North American mammalogy”. Genesis (2001).
29(3):105-109.
[28] Osgood WH. Revision of the mice of the American genus Peromyscus. North American fauna (1909).
28:1-285.
[29] Bedford NL, Hoekstra HE. Peromyscus mice as a model for studying natural variation. eLIFE (2015).
4:e06813.
[30] King JA. Biology of Peromyscus (Rodentia). American Society of Mammalogist (1968). Stillwater, Okla.
593p.
[31] Crossland JP, Dewey MJ, Barlow SC, Vrana PB, Felder MR, Szalai GJ. Caring for Peromyscus spp. in
research environments. Lab Anim (NY) (2014). 43(5):162-166.
[32] Steppan SJ, Adkins RM, Anderson J. Phylogeny and divergence-date estimates of rapid radiations of
muroid rodents based on multiple nuclear genes. Syst Biol (2004). 53(4):533-553.
[33] Joyner CP, Myrick LC, Crossland JP, Dawson WD. Deer mice as laboratory animals. ILAR Journal
(1998). 39(4):322-330.
[34] Foster DD. Differences in behavior and temperament between two races of the deer mouse. Journal of
Mammalogy (1959). 40(4):496-513.
[35] Weber JN, Peterson BK, Hoekstra HE. Discrete genetic modules are responsible for complex burrow
evolution in Peromyscus mice. Nature (2013). 493:402–405.
[36] Jašarevic E, Williams SA, Roberts RM, Geary DC, Rosenfeld CS. Spatial navigation strategies in
Peromyscus: a comparative study. Anim Behav (2012). 84(5):1141-1149.
[37] Dean P, Redgrave P, Lewis G. Locomotor activity of rats in open field after microinjection of procaine into
superior colliculus or underlying reticular formation. Behavioural Brain Research (1982). 5:175-187.
[38] Python Software Foundation. Python Language Reference, version 2.7. Available at
http://www.python.org.
[39] Mathis A, Mamidanna P, Cury KM, Abe T, Murthy VN, Mathis MW, Bethge M. DeepLabCut: markerless
pose estimation of user-defined body parts with deep learning. Nat. Neurosci (2018). 21.
[40] RStudio Team (2016). RStudio: Integrated Development for R. RStudio, Inc., Boston, MA URL
http://ww.rstudio.com/.
[41] Bradski G. The OpenCV Library, DDJ (2000).
[42] Zivkovic, Z. & Van Der Heijden, F. Efficient adaptive density estimation per image pixel for the task of
background subtraction. Pattern Recognit Lett (2006) 27:773–780.
[43] doi:10.5281/zenodo.1134016
[44] Tukey, J. W. (1977). Exploratory Data Analysis, Reading Massachusetts: Addison-Wesley.
[45] Krupa DJ, Ghazanfar AA, Nicolelis AL. Immediate thalamic sensory plasticity depends on corticothalamic
feedback. Proc Natl Acad Si USA (1999). 96:8200-8205.
[46] Soltani N, Mohammadi E, Allahtavakoli M, Shamsizadeh A, Roohbakhsh A, Haghparast A. Effects of
dimethyl sulfoxide on neuronal response characteristics in deep layer of rat barrel cortex. Basic and
Clinical Neuroscience (2016). 7(3):213-220.
[47] Hayes JP, Harrison RG. Variation in mitochondrial dna and the biogeographic history of woodrats
(neotoma) of the eastern united states. Syst. Biol (1992). 41(3):331–344.
[48] Runck AM, Cook JA. Postglacial expansion of the southern red-backed vole (Clethrionomys gapperi) in
North America. Mol Ecol (2005). 14:1445–1456.
[49] Riddle BR, Hafner DJ, Alexander LF. Phylogeography and systematics of the Peromyscus eremicus
species group and the historical biogeography of North American warm regional deserts. Mol Phylogenet
Evol (2000), 17(2):145–160.
[50] Sullivan J, Markert JA, Kilpatrick CW. Phylogeography and Molecular Systematics of the Peromyscus
Aztecus Species Group (Rodentia: Muridae) Inferred Using Parsimony and Likelihood. Syst Biol (1997).
46(3):426–440.
[51] Kalkvik HM, Stout IJ, Doonan TJ, Parkinson CL. Investigating niche and lineage diversification in widely
distributed taxa: Phylogeography and ecological niche modeling of the Peromyscus maniculatus species
group. Ecography (2012). 35:54–64.
[52] López-González C, Correa-Ramírez MM, García-Mendoza DF. Phylogeography of Peromyscus
schmidlyi : an endemic of the Sierra Madre Occidental, Journal of Mammalogy (2014). 95(2):254–26.








