
Ontwikkelingsstoornissen bestuderen met zebravissen
Wat is een ontwikkelingsstoornis? Het is een aandoening die vroeg in het leven optreedt en de normale ontwikkeling van een kind belemmert. Waarschijnlijk kent u wel iemand die er een heeft. Denk aan autisme, ADHD en intellectuele achterstand. Maar wat is de oorzaak van mentale ontwikkelingsstoornissen? Helaas is de oorzaak vaak niet gekend.
Aangezien er vaak niet veel geweten is over de oorzaak of het mechanisme van ontwikkelingsstoornissen, bestaan er maar een beperkt aantal therapieën voor deze aandoeningen. Verschillende onderzoeksgroepen focussen daarom hun aandacht op het zoeken naar oorzaken van ontwikkelingsstoornissen en op het begrijpen van de mechanismen die hierbij betrokken zijn. Door de mechanismen te begrijpen kunnen er nieuwe therapeutische doelwitten gevonden worden en nieuwe therapieën ontwikkeld worden.
16p11.2 deletiesyndroom
Zo werd er ontdekt dat een genetische fout, het 16p11.2 deletiesyndroom, kan leiden tot verschillende symptomen waaronder autisme, ADHD en intellectuele achterstand. Het 16p11.2 deletiesyndroom is een moeilijke naam voor een syndroom waarin een deel genetisch materiaal ontbreekt van chromosoom 16. Afhankelijk van hoe groot het deel is dat ontbreekt, krijg je andere symptomen.
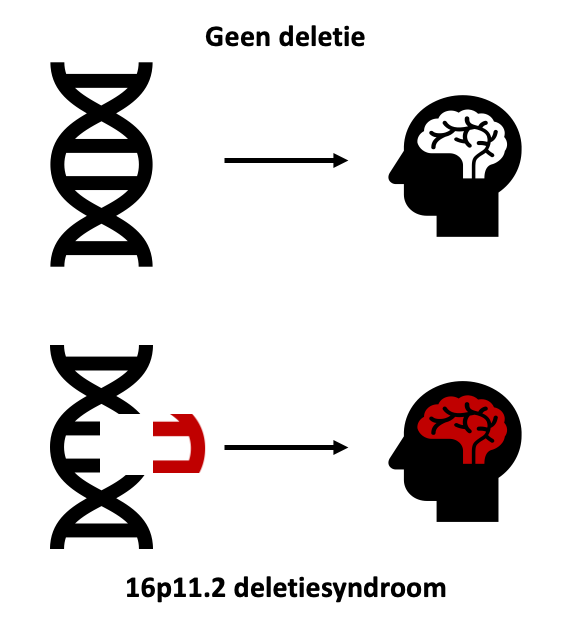
Figuur 1: Vereenvoudigde voorstelling van het 16p11.2 deletiesyndroom. Wanneer er geen deletie is, is de hersenfunctie normaal. In het 16p11.2 deletiesyndroom is er een deletie die leidt tot abnormale hersenfunctie.
Dus we weten dat het ontbreken van een deel van chromosoom 16 kan leiden tot verschillende ontwikkelingsstoornissen, maar hoe werkt dat nu precies?
Om dat te achterhalen is er onderzoek nodig. Chromosoom 16 bevat een groot aantal genen. Begrijpen wat die genen nu precies doen, hoe die samenwerken en wat er gebeurt als ze ontbreken, is cruciaal in het achterhalen van de werking van dat syndroom.
Zebravissen als onderzoeksmiddel
Dit is waar de vissen optreden. De genen die van belang zijn voor het 16p11.2 deletiesyndroom zijn ook aanwezig in zebravissen, de afwezigheid van die genen leidt ook tot symptomen in de zebravis. Dit is al een zeer belangrijke eigenschap voor het kiezen van een onderzoeksmodel. Daarbovenop is de ontwikkelingstijd van zebravissen relatief kort, ze reproduceren vaak en veel, wat het onderzoek ook vergemakkelijkt.
Zebravissen zijn sociale dieren. In de natuur blijven ze meestal in groep en reproduceren ze elkaars gedrag. Dit is een defensiemechanisme. Door in groep te blijven, zijn er meer ogen die op de uitkijk kunnen staan voor vijanden. Binnen de groep kunnen er natuurlijk ook ruzies ontstaan of in tegendeel romantische relaties. De zwempatronen van zebravissen zijn redelijk goed bestudeerd, er zijn bepaalde patronen die geassocieerd werden met agressief gedrag of paringsrituelen. Men kan ook achterhalen of een vis zich veilig voelt of niet, of die gestrest is of niet, op basis van zijn gedrag.
Dit alles leidt ertoe dat men zebravissen kan gebruiken om het 16p11.2 deletiesyndroom te bestuderen op genetisch, moleculair en gedragsvlak. Ze kunnen ons dus helpen met het begrijpen van het syndroom op verschillende niveaus.
Deletie van twee genen die belangrijk zijn voor het 16p11.2 deletiesyndroom
Om het 16p11.2 deletiesyndroom te begrijpen is het belangrijk om de functie van alle genen die hierbij betrokken zijn individueel te begrijpen en dan te kijken naar combinaties. Als we naar alle betrokken genen tegelijk zouden kijken, dan wordt het zeer ingewikkeld om een precies idee te krijgen van waar het precies mis gaat. We willen het syndroom in kleinere, makkelijker te onderzoeken deeltjes verdelen zodat we ons begrip van het syndroom geleidelijk aan kunnen opbouwen. Daarom hebben we ervoor gekozen om naar twee genen, fam57b en cdipt, te kijken. Deze twee genen zijn betrokken bij de vorming van een bepaald lipide (vet molecuul) dat belangrijk is voor de hersenen.
De hypothese is dat als deze twee genen niet normaal functioneren, de structuur van neuronen in de hersenen lichtjes verandert waardoor de hersenen abnormaal functioneren wat dan leidt tot afwijkende gedragspatronen.
Bevindingen op basis van het zwempatroon van vissen
We hebben het zwempatroon van normale vissen en genetisch gemanipuleerde vissen, voor de twee genen waarin we geïnteresseerd zijn, geanalyseerd in verschillende experimentele opstellingen. Hieruit hebben we verschillende conclusies kunnen trekken.
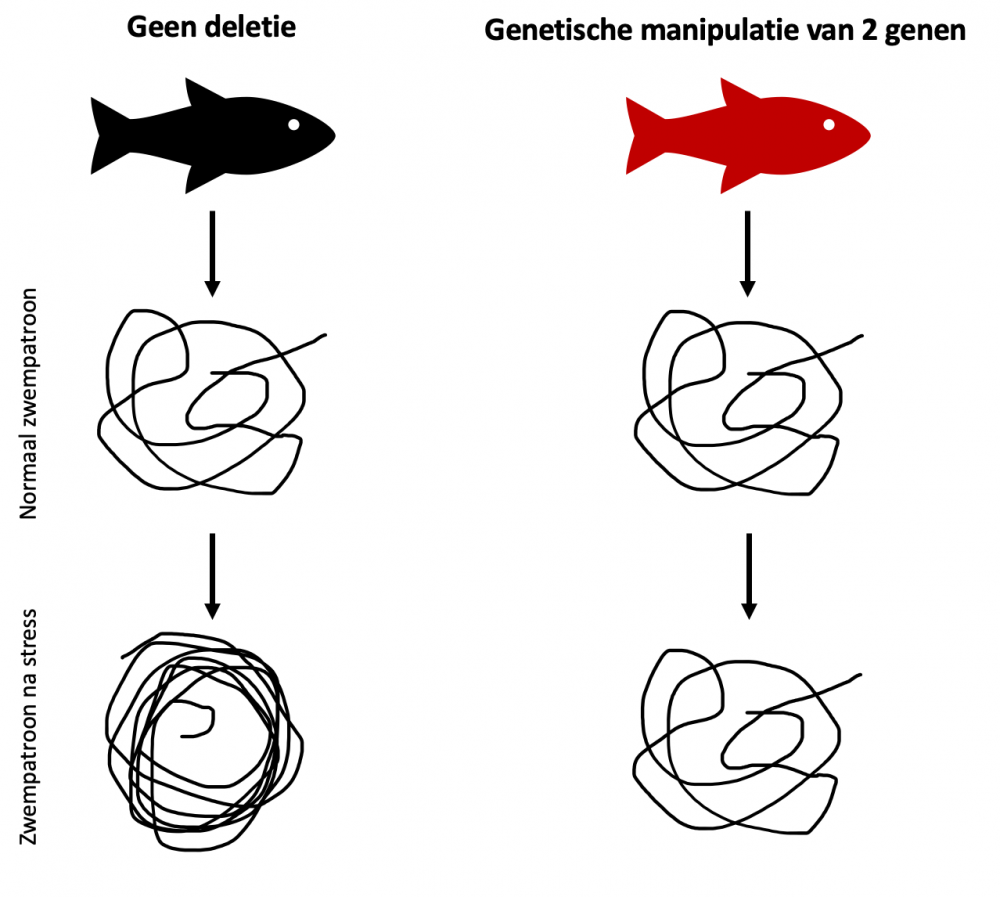
Figuur 2: Vereenvoudigde voorstelling van de analyse van het zwempatroon van vissen. Na blootstelling aan stress bewegen normale vissen meer dan onder gewone omstandigheden terwijl het zwempatroon van genetisch gemanipuleerde vissen niet significant verandert.
Door te kijken naar hoeveel afstand een vis aflegt voor en na blootstelling aan stress, kan men meer te weten komen over hoe gevoelig een vis is voor stress. Een andere manier om hiernaar te kijken, is door het verkennend gedrag van een vis te analyseren. Uit deze testen bleek dat onze genetisch gemanipuleerde vissen anders reageerden op stress dan onze normale vissen (controle vissen). De genetisch gemanipuleerde vissen lijken minder vatbaar te zijn voor stress.
We hebben ook gekeken naar de vatbaarheid voor epileptische aanvallen van de vissen na het toedienen van een bepaald geneesmiddel (een GABA-antagonist). Hieruit bleek dat de genetisch gemanipuleerde vissen vatbaarder waren voor beroertes dan de controle vissen. Dit betekent dus dat ze gevoeliger waren voor het geneesmiddel.
Recent werd er een artikel gepubliceerd dat aantoont dat stimulatie van een bepaalde soort neuronen (GABA-ergische inhibitorische neuronen) lijdt tot een vermindering van angstig gedrag. Aangezien onze genetisch gemanipuleerde vissen minder vatbaar lijken voor stress, dus minder angstig lijken en ze gevoeliger zijn voor een GABA-antagonist, concluderen we dat GABA-ergische inhibitorische neuronen een rol spelen in het verschil in gedrag dat we merken.
Conclusie
Dankzij deze studie hebben we geleerd en dat een bepaalde soort neuronen, namelijk de GABA-ergische inhibitorische neuronen, een rol spelen in het veroorzaken van de gedragsveranderingen van onze genetisch gemanipuleerde zebravissen. Dit brengt ons een klein stapje dichter tot het ontcijferen van de mechanismen die een rol spelen in het 16p11.2 deletiesyndroom. Er is nog veel onderzoek nodig vooraleer we aan therapeutische opties kunnen beginnen werken, maar elk onderzoek brengt ons iets dichter tot ons doel.
Bibliografie
-
Quality, C. f. B. H. S. a. 2015 National Survey on Drug Use and Health: Methodological summary and definitions. (Substance Abuse and Mental Health Services Administration, Rockville, MD, 2016).
-
Hanson, E. et al. The Cognitive and Behavioral Phenotype of the 16p11.2 Deletion in a Clinically Ascertained Population. Biological Psychiatry 77, 785-793, doi:10.1016/j.biopsych.2014.04.021 (2015).
-
Jacquemont, S. et al. Mirror extreme BMI phenotypes associated with gene dosage at the chromosome 16p11.2 locus. Nature 478, 97-U111, doi:10.1038/nature10406 (2011).
-
4 16p11.2 deletion syndrome, <https://ghr.nlm.nih.gov/condition/16p112-deletion-syndrome#statistics> (2019).
-
McCammon, J. M., Blaker-Lee, A., Chen, X. & Sive, H. The 16p11.2 homologs fam57ba and doc2a generate certain brain and body phenotypes. Human Molecular Genetics 26, 3699-3712, doi:10.1093/hmg/ddx255 (2017).
-
Pewzner-Jung, Y., Ben-Dor, S. & Futerman, A. H. When do lasses (longevity assurance genes) become CerS (ceramide synthases)? Insights into the regulation of ceramide synthesis. Journal of Biological Chemistry 281, 25001-25005, doi:10.1074/jbc.R600010200 (2006).
-
Yamashita-Sugahara, Y. et al. Fam57b ( Family with Sequence Similarity 57, Member B), a Novel Peroxisome Proliferator-activated Receptor gamma Target Gene That Regulates Adipogenesis through Ceramide Synthesis. Journal of Biological Chemistry 288, 4522-4537, doi:10.1074/jbc.M112.440792 (2013).
-
Cermenati, G. et al. Lipids in the nervous system: From biochemistry and molecular biology to patho-physiology. Biochimica Et Biophysica Acta-Molecular and Cell Biology of Lipids 1851, 51-60, doi:10.1016/j.bbalip.2014.08.011 (2015).
-
Hussain, G. et al. Role of cholesterol and sphingolipids in brain development and neurological diseases. Lipids in Health and Disease 18, doi:10.1186/s12944-019- 0965-z (2019).
-
Hannun, Y. A. & Obeid, L. M. The ceramide-centric universe of lipid-mediated cell regulation: Stress encounters of the lipid kind. Journal of Biological Chemistry 277, 25847-25850, doi:10.1074/jbc.R200008200 (2002).
-
Merrill, A. H. De novo sphingolipid biosynthesis: A necessary, but dangerous, pathway. Journal of Biological Chemistry 277, 25843-25846, doi:10.1074/jbc.R200009200 (2002).
-
Grösch, S., Schiffmann, S. & Geisslinger, G. Chain length-specific properties of ceramides. Prog Lipid Res 51, 50-62, doi:10.1016/j.plipres.2011.11.001 (2012).
-
Lykidis, A., Jackson, P. D., Rock, C. O. & Jackowski, S. The role of CDP- diacylglycerol synthetase and phosphatidylinositol synthase activity levels in the regulation of cellular phosphatidylinositol content. Journal of Biological Chemistry 272, 33402-33409, doi:10.1074/jbc.272.52.33402 (1997).
-
Hatch, G. M. & Vance, D. E. Stimulation of sphingomyelin biosynthesis by brefeldin A and sphingomyelin breakdown by okadaic acid treatment of rat hepatocytes. J Biol Chem 267, 12443-12451 (1992).
-
Pérez-Pérez, J. M., Candela, H. & Micol, J. L. Understanding synergy in genetic interactions. Trends in Genetics 25, 368-376, doi:10.1016/j.tig.2009.06.004 (2009).
-
Stewart, A. M., Braubach, O., Spitsbergen, J., Gerlai, R. & Kalueff, A. V. Zebrafish models for translational neuroscience research: from tank to bedside. Trends in Neurosciences 37, 264-278, doi:10.1016/j.tins.2014.02.011 (2014).
-
Blaker-Lee, A., Gupta, S., McCammon, J. M., De Rienzo, G. & Sive, H. Zebrafish homologs of genes within 16p11.2, a genomic region associated with brain disorders, are active during brain development, and include two deletion dosage sensor genes. 5, 834-851, doi:10.1242/dmm.009944 (2012).
-
Goldman, S. et al. Quantitative gait assessment in children with 16p11.2 syndrome. Journal of Neurodevelopmental Disorders 11, doi:10.1186/s11689-019-9286-9 (2019).
-
Rosenfeld, J. A. et al. Speech delays and behavioral problems are the predominant features in individuals with developmental delays and 16p11.2 microdeletions and microduplications. 2, 26-38, doi:10.1007/s11689-009-9037-4 (2010).
-
Panzini, C. M., Ehlinger, D. G., Alchahin, A. M., Guo, Y. P. & Commons, K. G. 16p11.2 deletion syndrome mice perseverate with active coping response to acute stress - rescue by blocking 5-HT2A receptors. Journal of Neurochemistry 143, 708- 721, doi:10.1111/jnc.14227 (2017).
-
Gillott, A. & Standen, P. J. Levels of anxiety and sources of stress in adults with autism. Journal of Intellectual Disabilities 11, 359-370, doi:10.1177/1744629507083585 (2007).
-
Muris, P., Steerneman, P., Merckelbach, H., Holdrinet, I. & Meesters, C. Comorbid Anxiety Symptoms in Children with Pervasive Developmental Disorders. 12, 387- 393, doi:10.1016/s0887-6185(98)00022-x (1998).
-
Van Steensel, F. J. A., Bögels, S. M. & Perrin, S. Anxiety Disorders in Children and Adolescents with Autistic Spectrum Disorders: A Meta-Analysis. Clinical Child and Family Psychology Review 14, 302-317, doi:10.1007/s10567-011-0097-0 (2011).
-
Lee, H. B. et al. Novel zebrafish behavioral assay to identify modifiers of the rapid, nongenomic stress response. Genes, Brain and Behavior 18, e12549, doi:10.1111/gbb.12549 (2019).
-
Bai, Y., Liu, H., Huang, B., Wagle, M. & Guo, S. Identification of environmental stressors and validation of light preference as a measure of anxiety in larval zebrafish. BMC Neuroscience 17, doi:10.1186/s12868-016-0298-z (2016).
-
Kalueff, A. V. et al. Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 10, 70-86, doi:10.1089/zeb.2012.0861 (2013).
-
Shinawi, M. et al. Recurrent reciprocal 16p11.2 rearrangements associated with global developmental delay, behavioural problems, dysmorphism, epilepsy, and abnormal head size. J Med Genet 47, 332-341, doi:10.1136/jmg.2009.073015 (2010).
-
Zufferey, F. et al. A 600 kb deletion syndrome at 16p11.2 leads to energy imbalance and neuropsychiatric disorders. Journal of Medical Genetics 49, 660-668, doi:10.1136/jmedgenet-2012-101203 (2012).
-
Baraban, S. C., Taylor, M. R., Castro, P. A. & Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience 131, 759-768, doi:10.1016/j.neuroscience.2004.11.031 (2005).
-
Berghmans, S., Hunt, J., Roach, A. & Goldsmith, P. Zebrafish offer the potential for a primary screen to identify a wide variety of potential anticonvulsants. Epilepsy Research 75, 18-28, doi:10.1016/j.eplepsyres.2007.03.015 (2007).
-
Thapar, A. & Cooper, M. Attention deficit hyperactivity disorder. The Lancet 387, 1240-1250, doi:10.1016/s0140-6736(15)00238-x (2016).
-
Lai, M.-C., Lombardo, M. V. & Baron-Cohen, S. Autism. The Lancet 383, 896-910, doi:10.1016/s0140-6736(13)61539-1 (2014).
-
Kalueff, A. V., Stewart, A. M. & Gerlai, R. Zebrafish as an emerging model for studying complex brain disorders. Trends in Pharmacological Sciences 35, 63-75, doi:10.1016/j.tips.2013.12.002 (2014).
-
Dreosti, E., Lopes, G., Kampff, A. R. & Wilson, S. W. Development of social behavior in young zebrafish. Frontiers in Neural Circuits 9, doi:10.3389/fncir.2015.00039 (2015).
-
Patowary, A. et al. Family-based exome sequencing and case-control analysis implicate CEP41 as an ASD gene. Translational Psychiatry 9, doi:10.1038/s41398- 018-0343-z (2019).
-
Ariyasiri, K. et al. Pharmacological (ethanol) and mutation (sam2 KO) induced impairment of novelty preference in zebrafish quantified using a new three-chamber social choice task. Progress in Neuro-Psychopharmacology and Biological Psychiatry 88, 53-65, doi:10.1016/j.pnpbp.2018.06.009 (2019).
-
Liu, C.-X. et al. CRISPR/Cas9-induced shank3b mutant zebrafish display autism-like behaviors. Molecular Autism 9, doi:10.1186/s13229-018-0204-x (2018).
-
Kurta, A. & Palestis, B. G. Effects of Ethanol on the Shoaling Behavior of Zebrafish (Danio Rerio). Dose-Response 8, dose-response.1, doi:10.2203/dose-response.10- 008.palestis (2010).
-
Sterling, M. E., Karatayev, O., Chang, G. Q., Algava, D. B. & Leibowitz, S. F. Model of voluntary ethanol intake in zebrafish: Effect on behavior and hypothalamic orexigenic peptides. 278, 29-39, doi:10.1016/j.bbr.2014.09.024 (2015).
-
Friard, O. & Gamba, M. BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods in Ecology and Evolution 7, 1325-1330, doi:10.1111/2041-210x.12584 (2016).
-
Baker, M. R., Goodman, A. C., Santo, J. B. & Wong, R. Y. Repeatability and reliability of exploratory behavior in proactive and reactive zebrafish, Danio rerio. Scientific Reports 8, doi:10.1038/s41598-018-30630-3 (2018).
-
Tea, J., Alderman, S. L. & Gilmour, K. M. Social stress increases plasma cortisol and reduces forebrain cell proliferation in subordinate male zebrafish (Danio rerio). The Journal of Experimental Biology 222, jeb194894, doi:10.1242/jeb.194894 (2019).
-
Schnörr, S. J., Steenbergen, P. J., Richardson, M. K. & Champagne, D. L. Measuring thigmotaxis in larval zebrafish. Behavioural Brain Research 228, 367-374, doi:10.1016/j.bbr.2011.12.016 (2012).
-
Norton, W. & Bally-Cuif, L. Adult zebrafish as a model organism for behavioural genetics. BMC Neuroscience 11, 90, doi:10.1186/1471-2202-11-90 (2010).
-
Assad, N. et al. Acute Restraint Stress Evokes Anxiety-Like Behavior Mediated by Telencephalic Inactivation and GabAergic Dysfunction in Zebrafish Brains. Scientific Reports 10, doi:10.1038/s41598-020-62077-w (2020).
-
Casillas-Espinosa, P. M., Powell, K. L. & O’Brien, T. J. Regulators of synaptic transmission: Roles in the pathogenesis and treatment of epilepsy. 53, 41-58, doi:10.1111/epi.12034 (2012).
-
Leweke, F. M., Louvel, J., Rausche, G. & Heinemann, U. Effects of pentetrazol on neuronal activity and on extracellular calcium concentration in rat hippocampal slices. 6, 187-198, doi:10.1016/0920-1211(90)90072-4 (1990).
-
Fukata, Y. & Fukata, M. Epilepsy and synaptic proteins. Curr Opin Neurobiol 45, 1- 8, doi:10.1016/j.conb.2017.02.001 (2017).
-
Lambert, C. J. et al. An automated system for rapid cellular extraction from live zebrafish embryos and larvae: Development and application to genotyping. PLOS ONE 13, e0193180, doi:10.1371/journal.pone.0193180 (2018).
-
Espinoza, G. J., Poland, J. M. & Bremer, J. R. A. Genotyping live fish larvae: Non-lethal and noninvasive DNA isolation from 3-5 day old hatchlings. Biotechniques 63, 181-186, doi:10.2144/000114598 (2017).
-
Wilkinson, R. N., Elworthy, S., Ingham, P. W. & van Eeden, F. J. M. A method for high-throughput PCR-based genotyping of larval zebrafish tail biopsies. Biotechniques 55, 314-316, doi:10.2144/000114116 (2013).
-
Kosuta, C. et al. High-throughput DNA Extraction and Genotyping of 3dpf Zebrafish Larvae by Fin Clipping. J Vis Exp, 58024, doi:10.3791/58024 (2018).






